El silicio (Si) es, tras el oxígeno, el elemento químico más abundante del planeta, forma parte de numerosas rocas y minerales, así como del manto terrestre. A nivel industrial, el silicio es importantísimo. Sus compuestos no solo son la base de numerosos adhesivos (siliconas, etc.) y materiales de construcción (fibras de vidrio, etc.), sino que también, por sus propiedades semiconductoras constituye el principal elemento de los microcircuitos de ordenadores y microchips de teléfonos móviles y otros numerosos aparatos. Asimismo, el silicio es el componente fundamental de las fibras ópticas que interconectan a las sociedades modernas.
Un escaso número de organismos —en su mayoría unicelulares, como diatomeas, algunos radiolarios, tecamebas, etc.— emplean silicio para elaborar sus esqueletos. Para ello, incorporan ácido silícico (Si OH4), un compuesto comúnmente presente en forma disuelta en el agua de mares, ríos y del suelo y, mediante diferentes procesos biológicos, lo polimerizan para formar sílice (SiO2). Dado que la fórmula química general de la sílice producida por procesos biológicos es la misma que la de la sílice del cristal de nuestras ventanas, puede decirse que los organismos silicificantes producen “esqueletos de cristal”. Entre los animales, el uso biológico del silicio es una rareza y solo es empleado por algunas especies y en concentraciones traza para diversas funciones. No obstante, existe un grupo de animales que constituyen una notable excepción: las esponjas.
Las esponjas son los animales vivos más antiguos y primitivos. Aparecieron en los mares hace más de 600 millones de años y son capaces de elaborar esqueletos formados exclusivamente por piezas de sílice. Estas piezas esqueléticas, que se denominan espículas, suelen medir solo unas micras o milímetros y aparecer en el cuerpo de la esponja en número de cientos de miles. Por el contrario, en algunas especies de aguas profundas, hay espículas que pueden medir hasta 3 m de longitud y, según dataciones isotópicas, su formación requiere unos 17 000 años.
Comprender los mecanismos celulares y moleculares que permiten a las esponjas convertir el silicio disuelto en el mar en estructuras complejas de sílice tiene un enorme interés científico. No solo porque puede ayudar a entender la evolución esquelética de los primeros animales, sino también por sus posibles aplicaciones para mejorar la producción biotecnológica de materiales silíceos.
Nuestro descubrimiento de que diferentes clases de esponja han desarrollado por evolución diferentes tipos de proteínas para polimerizar el ácido silícico y formar sus piezas esqueléticas de sílice tiene varias implicaciones científicas importantes:
1) En la actualidad, existen tres clases taxonómicas de esponjas con esqueletos silíceos, denominadas demosponjas, hexactinélidas y homoscleromorfas. Puesto que todas ellas poseen piezas esqueléticas de sílice, se había asumido tradicionalmente que empleaban mecanismos celulares y moleculares similares para construir sus esqueletos. El descubrimiento de que cada clase de esponja ha desarrollado su propio sistema de proteínas para polimerizar el ácido silícico revela que los esqueletos de sílice de las diferentes clases no son estructuras homólogas, sino “adquisiciones” evolutivas independientes. Es decir, cada clase de esponjas “ha inventado” por separado su propio sistema para fabricar un esqueleto silíceo.
2) Este hallazgo también revela que las esponjas ancestrales primero divergieron a nivel molecular y celular formando las tres clases y, solo después, cada una de las clases adquirió independientemente la capacidad de formar esqueletos de sílice. Si la capacidad de producir sílice se hubiera adquirido antes de que las clases de esponjas divergieran, todas las clases compartirían el mismo sistema de proteínas para la formación de sílice. Esta deducción es, en realidad, muy importante porque revela que tanto las esponjas ancestrales como los primeros representantes de las diferentes clases de esponjas eran organismos que carecían de esqueleto. Dicho escenario sugiere que, probablemente, el ancestro común de todos los animales también carecía de esqueleto. Los esqueletos aparecieron muchos millones de años después de la existencia de los primeros animales y de las primeras esponjas. En consecuencia, los paleontólogos deberían de reorientar y reconsiderar su búsqueda de fósiles con esqueleto mineral para datar el origen de los animales, puesto que la existencia de dichos esqueletos minerales es improbable.
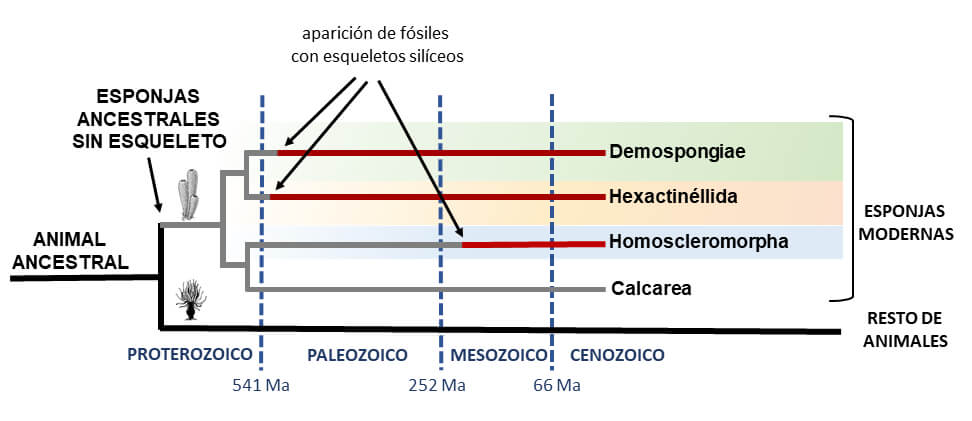
3) Hasta la fecha, se pensaba que existía un conflicto irresoluble entre las dataciones del origen de las esponjas mediante técnicas de relojes moleculares, que sitúan la aparición de las esponjas en el Proterozoico (hace unos 900-635 millones de años) y los restos esqueléticos fósiles, que indican un origen en el Cámbrico (hace unos 520 millones de años). Nuestros hallazgos indican que ambos métodos inferencias son correctos y compatibles: las esponjas aparecieron en el Proterozoico (como sugieren los relojes moleculares), pero carecían de esqueleto y, por tanto, no pudieron dejaron restos fósiles minerales hasta el Cámbrico (como atestigua el registro fósil).
El reto de la extracción e identificación de proteínas formadoras de sílice
La identificación de proteínas relacionadas con el proceso de formación de sílice que ha permitido hacer inferencias científicas sobre la evolución esquelética de los animales ancestrales constituye un reto científico importante, tanto por su dificultad técnica como por la necesidad de interpretar los resultados en un contexto multidisciplinar, que requiere conocimientos de biología molecular, biología celular, ecobiología de esponjas y de otros organismos silicificantes. Hasta la fecha, se había descrito una primera proteína en 1998, denominada silicateina, y, en 2015, una segunda proteína, denominada glasina. En nuestro estudio, se han identificado dos nuevas proteínas, hexaxilina y perisilina. Estas proteínas se han obtenido de esponjas de aguas profundas, cuyo estudio y recolección han sido posibles gracias al empleo de robots submarinos de alta tecnología.

El proceso empleado para la identificación de estas nuevas proteínas consistió primero en obtener las piezas esqueléticas de las esponjas. Para ello, se sumergieron muestras de tejido del cuerpo de los animales en lejía concentrada durante varios días y posteriormente en una mezcla de ácido nítrico y sulfúrico de alta concentración. Estos procesos digieren todos los componentes orgánicos del animal, quedando solo las piezas esqueléticas minerales limpias. Entonces, ¿cómo se pudieron encontrar proteínas silicificantes?
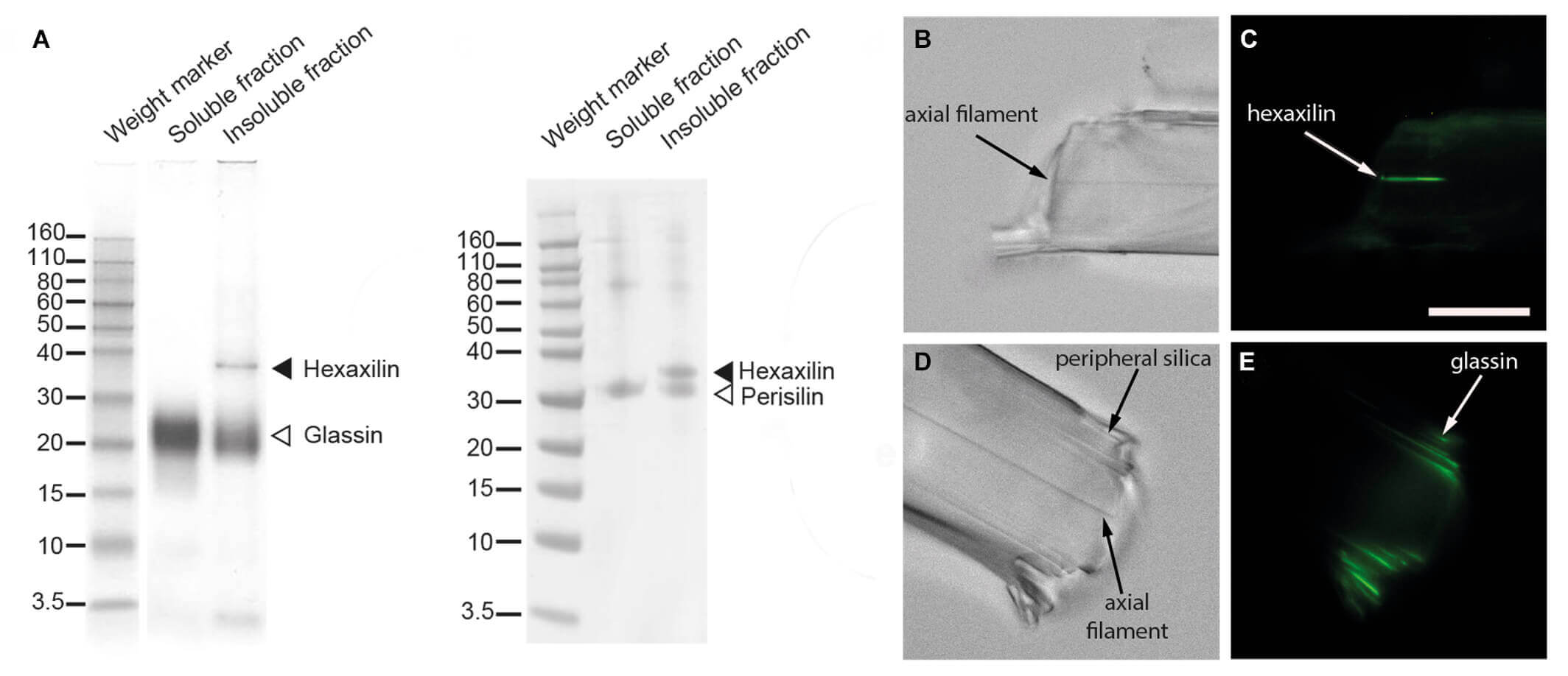
Las proteínas silicificantes están embebidas en el interior de la sílice de las piezas esqueléticas, por ello son las únicas que quedan protegidas del ataque de ácidos y bases durante el proceso de digestión química de los tejidos. Una vez las piezas esqueléticas estuvieron limpias, se procedió a disolver su sílice, es decir, la estructura de cristal, mediante inmersión en ácido fluorhídrico. Al disolverse el “cristal”, los componentes orgánicos que estaban embebidos en su interior quedan libres, bien como compuestos en disolución o bien como material no soluble. Estos restos solubles y no solubles se recolectaron y se hicieron migrar por un gel poliacrilamida de electroforesis, el cual permite separar las diferentes proteínas que pudieran estar presentes en la fracción soluble e insoluble, según su peso molecular. Como resultado, las proteínas con diferentes pesos moleculares forman bandas diferentes en el gel (ver figura). Una vez que los geles de electroforesis producen bandas que revelan la presencia de proteínas, es necesario identificar de qué proteínas se trata.
Para su identificación, las diferentes proteínas extraídas de la sílice se sometieron a un proceso de degradación controlada, ideado por Pehr V. Edman en 1950 y que, en su momento, constituyó un hito histórico. El proceso consiste en hacer reaccionar la proteína obtenida con fenilisotiocianato, de modo que el proceso marca solo el primer aminoácido del extremo N-terminal de la proteína y este se separa por hidrólisis ácida. El proceso se repite con segundo aminoácido, que ahora se ha convertido en el primero, y así sucesivamente. Los aminoácidos extraídos se identifican posteriormente mediante HPLC o, más modernamente, mediante secuenciadores. En nuestro caso, no se identificaron todos los aminoácidos de la proteína mediante la técnica de degradación de Edman, sino solo los 11 o 12 primeros del extremo N-terminal. Esta pequeña secuencia se empleó como referencia para identificar, mediante técnicas de bioinformática, la secuencia completa de la proteína en genomas y/o transcriptomas que se habían obtenido previamente para las especies de interés.
Una vez obtenida la secuencia completa de las nuevas proteínas, se seleccionó una pequeña zona de cada proteína (un péptido de 19 aminoácidos), que se sintetizó en laboratorio y fue inyectado en un conejo, para que este produjera un anticuerpo contra dicho péptido. El proceso de producción de un anticuerpo por el conejo requiere entre 5 y 6 meses. Finalmente, el anticuerpo se extrae del conejo en forma de suero, se purifica y, en nuestro caso, se asoció a un componente fluorescente (Alexafluor-488), que produce una emisión de luz verde cuando se ilumina con luz de una determinada longitud de onda al microscopio de fluorescencia.
Como puede observarse, la realización del estudio es un proceso largo y costoso que se prolongó durante cinco años y que requirió fondos de varios proyectos de investigación financiados por España y Japón, así como por el Programa H2020 de la Unión Europea.
Funcionamiento de las proteínas
La obtención de un anticuerpo que permitiera visualizar la localización de las nuevas proteínas identificadas resultó crucial para comenzar a entender cómo estas funcionan.
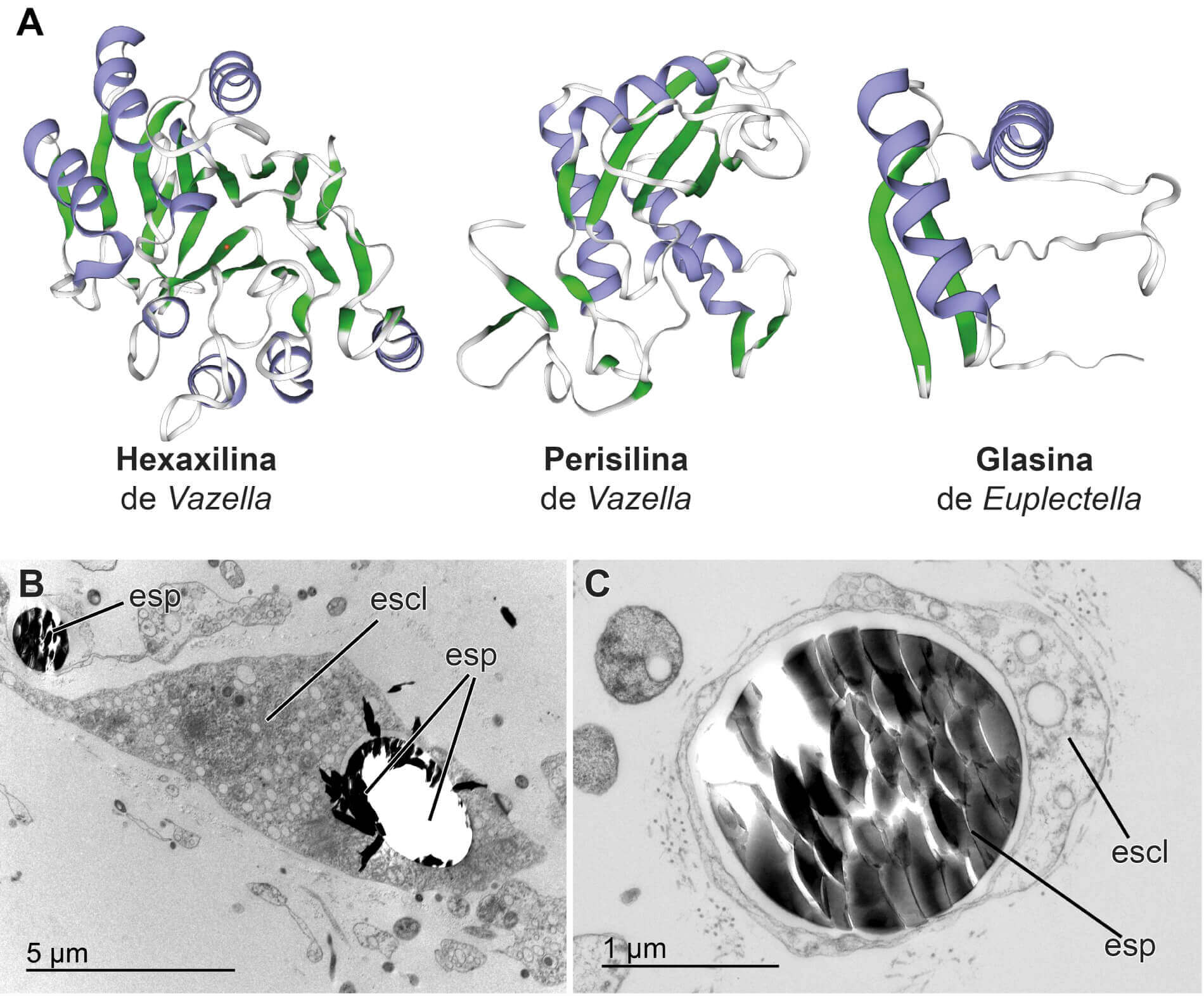
Se corroboró que la proteína hexaxilina aparece solo en la médula central de la pieza esquelética, mientras que la proteína perisilina aparece solo en la sílice periférica. Esto demostró que el proceso de polimerización del ácido silícico en sílice varía en función de si se está formando la parte interna o la parte periférica de la pieza esquelética.
Estos descubrimientos, junto con observaciones previas de microscopía electrónica, sugieren que las primeras fases de formación de espículas ocurren dentro de células especializadas, denominadas esclerocitos (ver figura). En el interior de los esclerocitos, se produciría la hexaxilina, que actuaría induciendo la deposición de sílice a su alrededor, hasta quedar finalmente embebida en la médula de la pieza esquelética. Posteriormente, las espículas serían liberadas por las células y, una vez en el medio extracelular, la perisilina se depositaría formando capas concéntricas, para conseguir el engrosamiento y la ornamentación final de la pieza esquelética. La formación de las piezas esqueléticas consta, por tanto, de varias fases y es un proceso más complejo de lo que se había asumido tradicionalmente.
A partir de la secuencia de aminoácidos de las diferentes proteínas silicificantes, se realizó una predicción de cómo estas se podrían plegar para convertirse en estructuras tridimensionales funcionales (ver figura). Dicha predicción estructural se realizó empleando el software AlphaFold, cuyos desarrolladores (D. Hassbis, J. Jumper y D. Baker) fueron galardonados con el Premio Nobel de Química 2024 por la importancia de dicho trabajo. Los modelos tridimensionales facilitarán el desarrollo de futuros estudios para entender, a nivel atómico, cómo estas proteínas consiguen inducir la polimerización de la sílice. A medio y largo plazo, los resultados podrían ayudar a mejorar los actuales procesos industriales de producción de sílice con fines biotecnológicos.
Referencias
- Katsuhiko Shimizu, Michika Nishi, Yuto Sakate, Haruka Kawanami, Tomohiro Bito, Jiro Arima, Laia Leria, Manuel Maldonado. Silica-associated proteins from hexactinellid sponges support an alternative evolutionary scenario for biomineralization in Porifera. Nature Communications, 2024, 15:181. doi: 10.1038/s41467-023-44226-7

Manuel Maldonado Barahona
Profesor de Investigación CSIC / Dr. En Biología

Cortesía de Muy Interesante
Dejanos un comentario: